Способ получения ингибитора с м зависимой эндонуклеазы
Похожие патенты | МПК / Метки | Текст | Заявка | Код ссылки



Текст
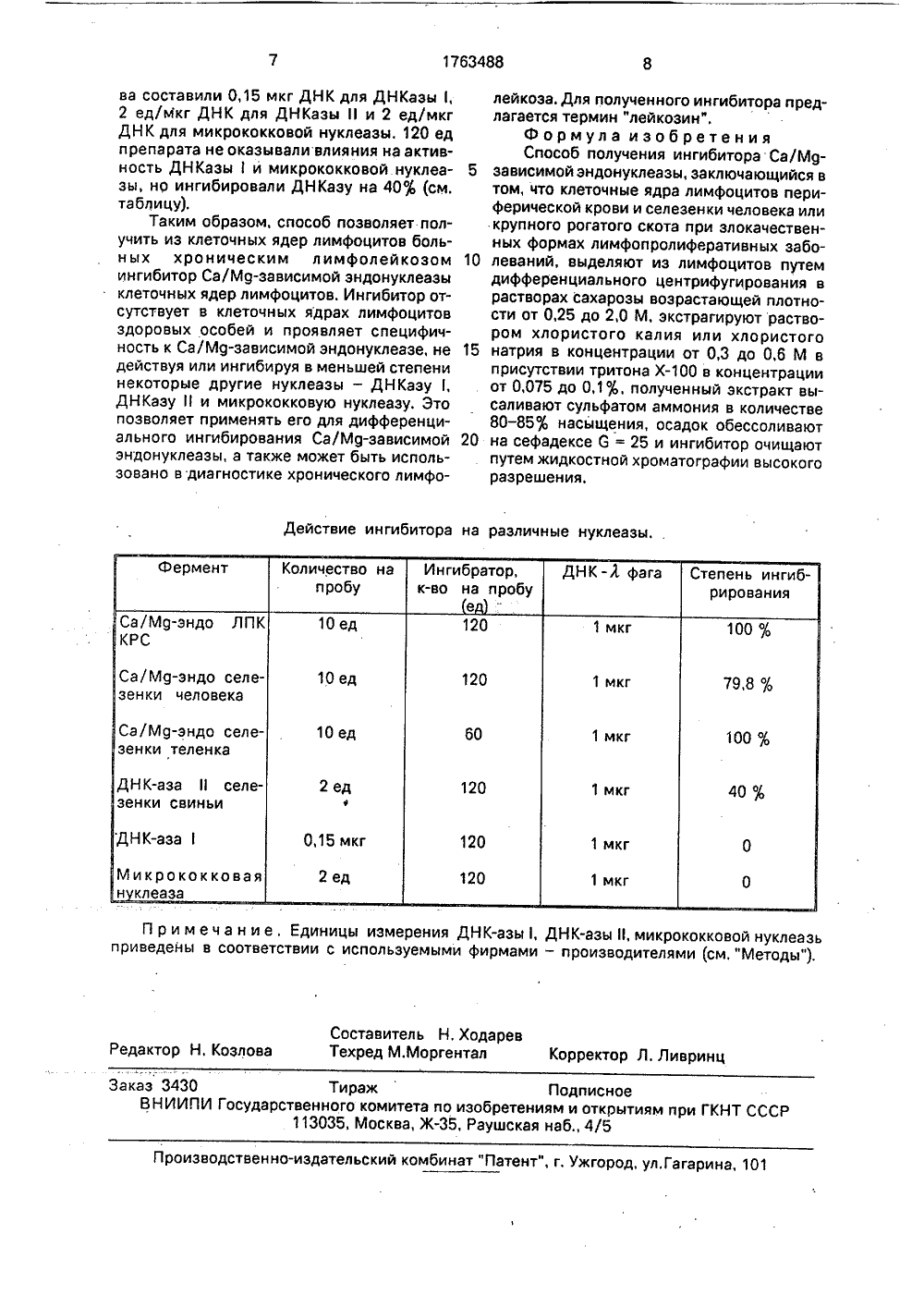
%35одственогииИ.В.Чупь С.С.Кирва и И.И ый центр меди ина, И,А.Сокоон, Ф.Е.Роман- отрин гибиторв литерае. Извесиальныхн ингибвусез е леазы вперв бакте получ Асуп о ГОСУДАРСТВЕННЫЙ КОМИТЕТПО ИЗОБРЕТЕНИЯМ И ОТКРЫТИЯМПРИ ГКНТ СССР ВТОРСКОМУ СВИДЕТЕЛЬ(57) Использование; биохимия, получениеингибитора Са/Мд-зависимой эндонуклеазы лимфоцитов. Сущность изобретения: вкачестве объекта выделения используют Изобретение относится к биохимии, в частности к получению ингибиторов нуклеаз.Ингибиторы нуклеаз могут быть использованы для дифференциального подавления активности отдельных ферментов в инкубационных смесях, содержащих несколько нуклеаз. Они также могут применяться для получения интактных препаратов нуклеиновых кислот и для аффинной хроматографии нуклеаз, Исследование ингибиторов нуклеаз важно для понимания механизмов регуляции этих ферментов в клетке. Са/Мд-зависимой эндонуктуре не известен и получен тны природные ингибиторы эндонуклеаз. В частности, итор внеклеточной ДНКазы чог 3 2789, Ингибитор выделимфоциты периферической крови или селезенки человека, или крупного рогатого скота при злокачественных лимфопролиферативных заболеваниях, наиболее предпочтительно - при хроническом лимфолейкозе, из лимфоцитов выделяют клеточные ядра в растворах сахарозы возрастающей плотности(от 0,25 до 2,0 М), клеточные ядра экстрагируют растворами 0,3-0,6 М ИаС или КС в присутствии 0,075-0,1 -ного тритона Х, экстракты высаливают сульфатом аммония в количестве 80-85; насыщения и из обессоленного осадка выделяют ингибитор с помощью высокоэффективной жидкостной колоночной хроматографии. ляли из экстрактов мицелия культур, фракционируя их сульфатом аммония и хроматографируя на сефадексе 6-100 и КМ-целлюлозе, Ингибитор был охарактеризован как лабильный белок с рЬ 7,8 - 8,2, специфичный к ДНКазе 1 А ечогз 2789 (1). О Известны также природные ингибиторы эн- Ы донуклеаз, полученные из тканей животных, фь Например, из селезенки телят выделен ин- СО гибитор ДНКазы 1 (панкреатической), Инги битор выделяли, фракционируя экстракт сульфатом аммония и хроматографируя активные фракции на геле окиси алюминия и ДЭАЭ-целлюлозе. Ингибитор был идентифицирован как 0-актин или близкородственный ему белок (2). Данный ингибитор не действует на Са/Мд-зависимую эндонуклеазу клеточных ядер, в частности из лимфоидных тканей, Природные ингибиторы Са/Мд-зависимой эндонуклеазы в литературе не описаны и нами получен впервые.5 10 15 20 25 30 35 40 45 50 55 Цель изобретения - получение природного ингибитора Са/Мц-зависимой эндонуклеазы клеточных ядер лимфоцитов.Поставленная цель достигается тем, что для получения ингибитора используютлимфоциты периферической крови или селезенки крупного рогатого скота или человека при злокачественных формах лимфопролиферативных заболеваний - наиболее предпочтительно при хроническом лимфолейкозе (ХЛЛ), причем ингибитор вы деляют не из целостных клеток, а из изолированных клеточных ядер, Для этого клеточные ядра экстрагируют раствором, содержащим 0,3 - 0,6 М КС с 0-0,1 -ным тритоном Х, экстракт фракционируют сульфатом аммония до 80 - 85 фнасыщения и активные фракции после растворения и диализа подвергают обессоливанию на колонках с 6-25 и высокоэффективной жидкостной колоночной хроматографии на катионообменных колонках "Мапо Я" (Фармация, Швеция), с элюцией градиентом хлористого натрия от 0 до 0,5 М. Способ позволяет получить препарат, обладающий выраженной ингибирующей активностью по отношению к Са/М 9-зависимой эндонуклеазе клеточных ядер лимфоцитов. Ингибирующая активность специфична, так как препарат не ингибирует ДНКазу 1 и микрококковую нуклеазу и связана с белком.Изобретение иллюстрируется следующими примерами.П р и м е р 1, Лимфоциты периферической крови, полученные от коровы, большой хроническим лимфолейкозом, суспендировали в растворе, содержащем 10 вМ трис- НС, рН 8,0, 3 вМ М 9 С 2, 0,9 М сахарозу и 0,002 тритона Х(ТМС,9); на 10 лимфоцитов использовали 10 мл среды. Суспензию гомогенизировали в гомогенизаторе Э вел ьгейма-Поттера и наслаивали на 10 пМ трис-НС, рН 8,03 вМ М 9 СЬ, 1,2 М сахарозу в соотношении 3:1. Пробы центрифугировали при 1500 - 20009 10 мин при 4 С. К осадкам добавляли 10 мл ТМС,9, тщательно суспендировали и повторно центрифугировали в том же режиме, Осадки были представлены интактными клеточными ядрами.Клеточные ядра суспендировали в 10 гпМ трис-НС, рН 7,4 - 7,6, до конечной концентрации 2 мг/мл (по ДНК). К суспензии добавляли 3 объема 0,4 М КС, 0,1 тритона Хв 10 гпМ трис-НС, рН 7,4 - 7,6(конечные концентрации КС и тритона Хсоставляли 0,3 М и 0,075 фсоответственно) и перемешивали на ледяной бане 30 - 60 мин. После этого пробы центрифугировали при 24000 д 15 мин, 4 С, Осадки отбрасывали, В надосадках (" экстрактах" ) содержались негистоновые белки хроматина и ингибирующие факторы. Специальные исследования относительно условий экстракции показали следующее. При конечной концентрации КС в экстрагирующей смеси 0,3 М (см. выше) степень экстракции ядерных белков составляет 21 ф(степень экстракции определяли по формуле (а/а+Ь)100, где "а" - количество белка в экстракте, "Ь" - количество белка в осадке). При снижении конечной концентрации КС (до 0,2 и 0,1 М) степень экстракции существенно снижалась, снижался и выход целевого продукта. При повышении концентрации КС до 1,0 М степень экстракции белков возрастала до 45-50%, однако активность ингибитора в экстракте не возрастала после концентрации 0,6 М. Введение в экстрагирующую смесь тритона Хв конечной концентрации 0,05-0,075 повышало степень экстракции белков в присутствии 0,3 М КС до 46 , при этом активность ингибитора возрастала на 20-25. Другие исследованные условия экстракции ядерных белков - ультразвуковая обработка ядер, экстракция гуанидинхлоридом в присутствии дитиотрейтола, применение только триона - Х- давали худшие результаты.Полученные экстракты фракционировали высаливанием сульфатом аммония. Для этого пробы обессоливали на сефадексе С и при непрерывном перемешивании добавляли сухой сульфат аммония до 80 насыщения. Сформировавшийся осадок отделяли центрифугированием при 100000 9, 1 ч, 4 С; надосадок отбрасывали. Применение высаливания сульфатом аммония обеспечивало концентрирование образцов, освобождение от части балластных белков и стабилизацию ингибитора. Полученные осадки могли храниться при 4 С без потери активности 1 месяц. Понижение концентрации сульфата аммония до 70 приводило к снижению выхода ингибитора на 10-15 и снижению стабильности препарата. Повышение концентрации сульфата аммония до 85 фи 90 фне приводило к изменению результатов,Осадок, полученный после высаливания, растворяли в минимальном объеме 10 вМ трис-НС, рН 7,4 - 7,6 и обессоливали на колонках с сефадексом 6-25. Обессоленный образец, содержащий 1 - 1,5 мг белка в 1-2 мл, наносили на катионообменную колонку "Мопо Я" (система РР С, Фармация, Швеция), предварительно уравновешенную тем же буфером. Скорость нанесения составляла 0,1-0,2 мл/мин. После нанесения образца колонку промывали 5 - 7 мл того же буфера со скоростью 1 мл/мин, после чегоначинали элюцию сорбированных белков линейным градиентом йаС от 0 до 0,5 М в том же буфере. Скорость элюции - 1 мл/мин, давление - 1,0-1,5 МРа, обьем градиента - 10 мл, Собирали фракции обьемом 0,5 или 1,0 мл и анализировали их на ингибиторную активность.Активность ингибитора определяли следующим образом. Готовили инкубационные пробы, содержащие 40 мкл 10 аМ трис- НС, рН 7,6, 10 аМ МдС 2, 1 тМ СаС 2, 10 п 1 М 2-меркаптоэтанол и 10 ед Са/Мц-зависимой эндонуклеазы клеточных ядер лимфоцитов здоровых животных. Пробы готовили в лунках 96-луночных микроплат. За единицу активности фермента принимали его количество, достаточное для полного расщепления 0,1 мкг ДНК Л-фага за 2 часа инкубации при 37 С в средах, содержащих 10 щМ МцС 2, 1 гпМ СаС 2. Полноту расщепления контролировали электрофорезом в гелях 0,8% агарозы,К инкубационным пробам добавляли 10 мкл каждой фракции градиента и инкубировали на льду 5 - 10 мин. Затем в пробы вносили 1 мкг ДНК Л -фага и инкубацию продолжали в течение 2 часов при 37 С. После этого в пробы вносили 5 мкл 50% глицерина с 2,50 бромфенолового синего и наносили их на горизонтальный блок геля 0,8% агарозы, Электрофорез проводили при напряжении 150 В; гели окрашивали бромистым этидием и фотографировали в отраженном ультрафиолетовом свете с длиной волны 254 нм,Во фракциях, соответствующих концентрации КаС от 0,25 до 0,3 М, обнаруживалось сохранение интактной (нерасщепленной) ДН К Л-фага, что говорит о присутствии в них ингибитора Са/Мц-зависимой эндонуклеазы.За единицу активности препарата ингибитора было принято такое его количество, которое обеспечивало ингибирование 10 ед Са/Мд-зависимой эндонуклеазы на 1%, Выход ингибитора составлял 12000 ед/мг ДНК исходных ядер, удельная активность - 600 ед/мкг белка, очистка - в 100 раз по отношению к экстракту,По данным электрофореза в полиакриламидных гелях сдодецилсульфатом натрия и обработок препарата различными протеазами, ингибирующая активность препарата связана с полипептидами размерами от 60 до 67 кО.П р и м е р 2, Все операции проводили аналогично примеру 1, за исключением того, что использовали лимфоциты периферической крови людей, больных хроническим лимфолейкозом, и активность ингибитораопределяли непосредственно к экстрактахклеточных ядер,П р и м е р 3. Для препаративного выделения ингибитора использовали селезенку5 КРС, больного спонтанным ХЛЛ,Все процедуры выполняли как описановыше, за исключением того, что клеточныеядра выделяли следующим образом, В опытбрали 0,2 кг селезенки, Образец измельчали10 в мясорубке. Полученную массу переносилив 5-литровую колбу и заливали 2 л охлажденного буфера А (0,90 йаС, 0,5% Иа-цитрат,10 мМ трис-НС, рН 7,6). Колбу переносилив баню со льдом и содержимое перемеши 15 вали, интенсивно вращая колбу 30 мин. Полученную суспензию фильтровали через 4слоя марли и фильтрат центрифугировалипри 5000 ц 30-40 мин. К осадку добавляли 1л буфера В (0,25 М сахарозы, 0,4% нонидет20 Р, 5 мМ СаС 2, 50 мМ трис-НС, рН 8,5) игомогенизировали порциями по 60 мл в гомогенизаторе Поттера. Гомогенат фильтровали через 4 слоя марли ицентрифугировали при 5000 ц 30 мин, По25 данным микроскопии осадок более, чем на900 состоял из лимфоцитов. К осадку добавляли 500 мл буфера С (1,0 М сахароза, 5мМ СаС, 50 мМ трис-НС, рН 8,5, 0,4% нонидет Р) и интенсивно гомогенизирова 30 ли. Гомогенат центрифугировали при 50000ц 30 мин. Осадки суспендировали в буфереС и наслаивали в 100-мл центрифужных пробирках на 20 мл буфера Е (2,0 М сахароза,50 мМ трис-НС, рН 8,5, 5 мМ СаС 2) и цент 35 рифугировали при 70000 ц 2 часа.П р и м е р 4. Для выделения ингибитораиспользовали клеточные ядра, полученныеиз лимфоцитов периферической крови здорового животного. Все этапы проводили40 аналогично примеру 1. Ингибирующей активности обнаружить не удалось.П р и м е р 5, Все приемы выполнялианалогично примеру 1, за исключением того, что определяли действие ингибитора на45 Са/Мд-зависимую эндонуклеазу клеточныхядер лимфоцитов селезки здоровых животных и человека. 120 ед препарата ингибировали активность фермента селезенкикрупного рогатого скота на 100%, а селезен 50 ки человека - на 800 .П р и м е р 6. Определяли действиеингибитора на коммерческие препаратыДНКазы(панкреатической), ДН Казы(селезенки свиньи) и микрококковой нуклеазы.55 Количества фермента подбирали так, чтобыэлектрофоретическая картина расщепленияДНК Л -фага была сходна с таковой дляиспользуемых количеств Са/Мд-зависимойэндонуклеазы. Соответствующие количест1763488 Действие ингибитора на различные нуклеазы. ДНК-А фага Ингибратор,к-во на пробуе Количество на пробуЗаказ 3430 Тираж Подписное ВНИИПИ Государственного комитета по изобретениям и открытиям при ГКНТ СССР 113035, Москва, Ж, Раушская наб., 4/5 Производственно-издательский комбинат "Патент", г. Ужгород, ул.Гагарина, 101 ва составили 0,15 мкг ДНК для ДНКазы 1, 2 ед/Мкг ДНК для ДНКазы 11 и 2 ед/мкг ДНК для микрококковой нуклеазы. 120 ед препарата не оказывали влияния на активность ДН Казыи микрококковой нуклеазы, нр ингибировали ДНКазу на 40 о(см. таблицу).Таким образом, способ позволяет получить из клеточных ядер лимфоцитов больных хроническим лимфолейкозом ингибитор Са/Мд-зависимой эндонуклеазы клеточных ядер лимфоцитов. Ингибитор отсутствует в клеточных ядрах лимфоцитов здоровых особей и проявляет специфичность к Са/Мд-зависимой эндонуклеазе, не действуя или ингибируя в меньшей степени некоторые другие нуклеазы - ДНКазу 1, ДНКазу И и микрококковую нуклеазу. Это позволяет применять его для дифференциального ингибирования Са/Мд-зависимой эндонуклеазы, а также может быть использовано в диагностике хронического лимфолейкоза. Для полученного ингибитора предлагается термин "лейкозин".Формула изобретения Способ получения ингибитора Са/Мд зависимой эндонуклеазы, заключающийся втом, что клеточные ядра лимфоцитов периферической крови и селезенки человека или крупного рогатого скота при злокачественных формах лимфопролиферативных забо леваний, выделяют из лимфоцитов путемдифференциального центрифугирования в растворах сахарозы возрастающей плотности от 0,25 до 2,0 М, экстрагируют раствором хлористого калия или хлористого 15 натрия в концентрации от 0,3 до 0,6 М вприсутствии тритона Хв концентрации от 0,075 до 0,1%, полученный экстракт высаливают сульфатом аммония в количестве 80 - 85% насыщения, осадок обессоливают 20 на сефадексе 0 = 25 и ингибитор очищаютпутем жидкостной хроматографии высокого разрешения.
СмотретьЗаявка
4911591, 28.12.1990
НАУЧНО-ПРОИЗВОДСТВЕННЫЙ ЦЕНТР МЕДИЦИНСКОЙ БИОТЕХНОЛОГИИ
ХОДАРЕВ НИКОЛАЙ НИКОЛАЕВИЧ, ЧУПЫРИНА ИННА ВАСИЛЬЕВНА, СОКОЛОВА ИРИНА АНАТОЛЬЕВНА, ТКАЧЕВА ТАТЬЯНА ИВАНОВНА, КИРЗОН СЕРАФИМА СЕМЕНОВНА, РОМАНЦЕВ ФЕДОР ЕВГЕНЬЕВИЧ, ПЧЕЛИНЦЕВА ОЛЬГА АЛЕКСАНДРОВНА, ВОТРИН ИГОРЬ ИВАНОВИЧ
МПК / Метки
МПК: C12N 9/16, G01N 27/26
Метки: зависимой, ингибитора, эндонуклеазы
Опубликовано: 23.09.1992
Код ссылки
<a href="https://patents.su/4-1763488-sposob-polucheniya-ingibitora-s-m-zavisimojj-ehndonukleazy.html" target="_blank" rel="follow" title="База патентов СССР">Способ получения ингибитора с м зависимой эндонуклеазы</a>
Способ получения биомассы

Номер патента: 578901
Опубликовано: 30.10.1977
Авторы: Патрик, Филип, Фрэнсиз
МПК: C12D 13/06
Метки: биомассы
...разбавлениядолжна быть значительно ниже ожидаемой максимальной скорости роста микроорганизмов. Оптимальная рабочая температура более 40 С, скорость разбавления О,е 5-0,15 ч.Пример.1. Стадия подготовки культуры.500 мл шлама, содержащего метан грязи из отстойника, добавляют с интервалами в течение 5 дней в аэрируемый ферментатор с 5 л жидкой питательной среды следующего состава, мг/л:КН 1 О 1600фОНР 1 1, 12 НО 2928Ма МО 1180Мпво, 7 Н,О 80РеЬО, 7 НО 14Со(МО) 1 11 НО 25СаЬО 5 Н,О4гкьо 17 НО 0,34дпЯцН 20 О,3 ОйаМО, 2 НрО 0,24Дистиллйрованной водой доводято объем до 1 л, Температура 45 С, скорость разбавления 0,084 ч , скорость мешалки Ферментатора 1000 об/мин, рН 6,75 создают добавлением 0,5 н. раствора едкого натра.В...
Способ оценки степени повреждения головного мозга

Номер патента: 1686362
Опубликовано: 23.10.1991
Авторы: Григорьев, Кашина, Паничев
МПК: A61B 10/00, G01N 33/50
Метки: головного, мозга, оценки, повреждения, степени
...Данных за компрессию головного мозга нет, Таким образом данные о повреждении головного мозга, полученные на основании определения содержания меди, совпадают с клиническим диагнозом, Небольшая скорость снижения концентрации меди 2/мин могла указывать на незначительное кровоизлияние, что впоследствии было подтверждено данными патологоанатомического исследования, выявившего наряду с очагами раэмозжения в височных долях, небольшое "сателлитное" субдуральное кровоизлияние в левой лобновисочной области.П р и м е р 5, Больной С 25 лет, Поступил через 40 мин после драки, В соответствии с предлагаемым способом сразу же взята кровь в количестве 1 мл из локтевой вены и определена концентрация меди, которая составила 1,8 мкг/мл, Этот результат...
Штамм гибридных культивируемых клеток животных mus мusсulus продуцент моноклональных антител к са м зависимой эндонуклеазе клеточных ядер лимфоцитов селезенки человека

Номер патента: 1497213
Опубликовано: 30.07.1989
Авторы: Волгин, Волгина, Вотрин, Певницкий, Ходарев
МПК: A61K 39/395, C12N 5/00
Метки: антител, гибридных, животных, зависимой, клеток, клеточных, культивируемых, лимфоцитов, мusсulus, моноклональных, продуцент, селезенки, человека, штамм, эндонуклеазе, ядер
...скота,Для выращивания штамма используютстеклянную или пластиковую посуду,Во Флакон объемом 25 см в 5 мл средывносят 2 10 -10 клеток РИ, Пассажи б1 раз в 3-4 дня той же дозой клеток.Кратность рассева 1:4.Культивирование в организме животного.Для выращивания опухоли из штаммапригодны мыши линии ВАЬВ/с. Свежиммьппам за 7-30 (оптимально 14) днейдо инъекции штамма вводят внутрибрюшинно 0 5 мл пристана, Штамм инъециЭ6руют внутрибрюшинно по 3,2 х 10 клеток на мышь в 1 мл среды Игла. Асцитическую жидкость собирают по мерепоявления (на 10-15 день). Из асциташтамм перевивают на свежих мышей по1-5 х 10 клеток на мышь, 4-6 пассажей, 14972136Продуктивность штамма,Секреция монАТ на 4-й день культивирования 10-20 мкг в 1 мл среды и1-2 мг в 1 мл...
Штамм гибридных культивируемых клеток животных mus мusсulus продуцент моноклональных антител к са mg зависимой эндонуклеазе клеточных ядер лимфоцитов селезенки человека

Номер патента: 1497214
Опубликовано: 30.07.1989
Авторы: Волгин, Волгина, Вотрин, Певницкий, Ходарев
МПК: A61K 39/395, C12N 5/00
Метки: антител, гибридных, животных, зависимой, клеток, клеточных, культивируемых, лимфоцитов, мusсulus, моноклональных, продуцент, селезенки, человека, штамм, эндонуклеазе, ядер
...иммунизации - 13 позитивных клонов из 200 первичных культур, Бсе стабильные позитивные линииклонированы и заморожены в жидкомазоте. Клон РЯ клонирован еще 5 раз.Для наработки антител клон Ы культивируют в условиях хп ч 1 чо в видеасцитной жидкости в брюшной полостисингенных мышей,Штамм депонирован под номеромВСКК (П) Р 117 Д и характеризуетсяследующими признаками.Культуральные признаки,Стандартные условия культивирования, Среда для культивирования - среда Дульбеко с добавлением 107. телячьей эмбриональной сыворотки., 4 ммЬ-глутамина. 80 мкг/мл гентамицинаОУ37 С. 57. СО, 967. влажности. Допустимо использование среды Игла.с добавлением 157 сыворотки крупного рогатого скота.Для выращивания штамма используютстеклянную или пластиковую...
Способ определения активности с м -зависимой эндонуклеазы лимфоцитов

Номер патента: 1541514
Опубликовано: 07.02.1990
Авторы: Александрова, Вотрин, Газдаров, Коромыслов, Ходарев, Чупырина
МПК: G01N 33/48
Метки: активности, зависимой, лимфоцитов, эндонуклеазы
...3 ммольМрСЭ.; 1,2 моль сахарозы) в соотноше -нии 1:3 и центрифугируют 1 О мин,1500-2000 р, при 4 Г,Осадки ядер суспендируют в минимальном объеме ТМС,9 беэ тритонаХ, затем доводят объем ТМС,9до 10-12 мл и суспенэию центрифугируют 1500-2000 я при 4 С 10 мин. Осадки поданным световоймикроскопии представляют собои интактные клеточныеядра, свободные от цитоплаэматических загрязнений, Выход ядер 0,3 мгДНК/10 лимфоцитов,8Для определения активности Са/Мрзависимой зндонуклеазы клеточные ядра суспендируют в инкубационной средеТМСС (50 ммоль трис-НС 1, рН 8,3;10 ммоль МрС:.2, 1 ммоль СаС 1;0,25 моль сахарозы) до концентрации1 мг/мл по ДНК, В инкубационные пробирки (.Эппендорф) вносят 10-50 мклсуспензии ядер в ТМСС. Инкубацию проводят при 37 С в...
Предыдущий патент: Способ определения дезадаптации населения к условиям окружающей среды
Следующий патент: Устройство для регулирования рн и парциального давления растворенного кислорода в культуральной жидкости в ферментере
Случайный патент: Устройство для отливки постоянных магнитов, состоящее из обогреваемой формы и расположенного под ней охлаждаемого кристаллизатора