Способ определения моноаддуктов и диаддуктов днк
Похожие патенты | МПК / Метки | Текст | Заявка | Код ссылки



Текст
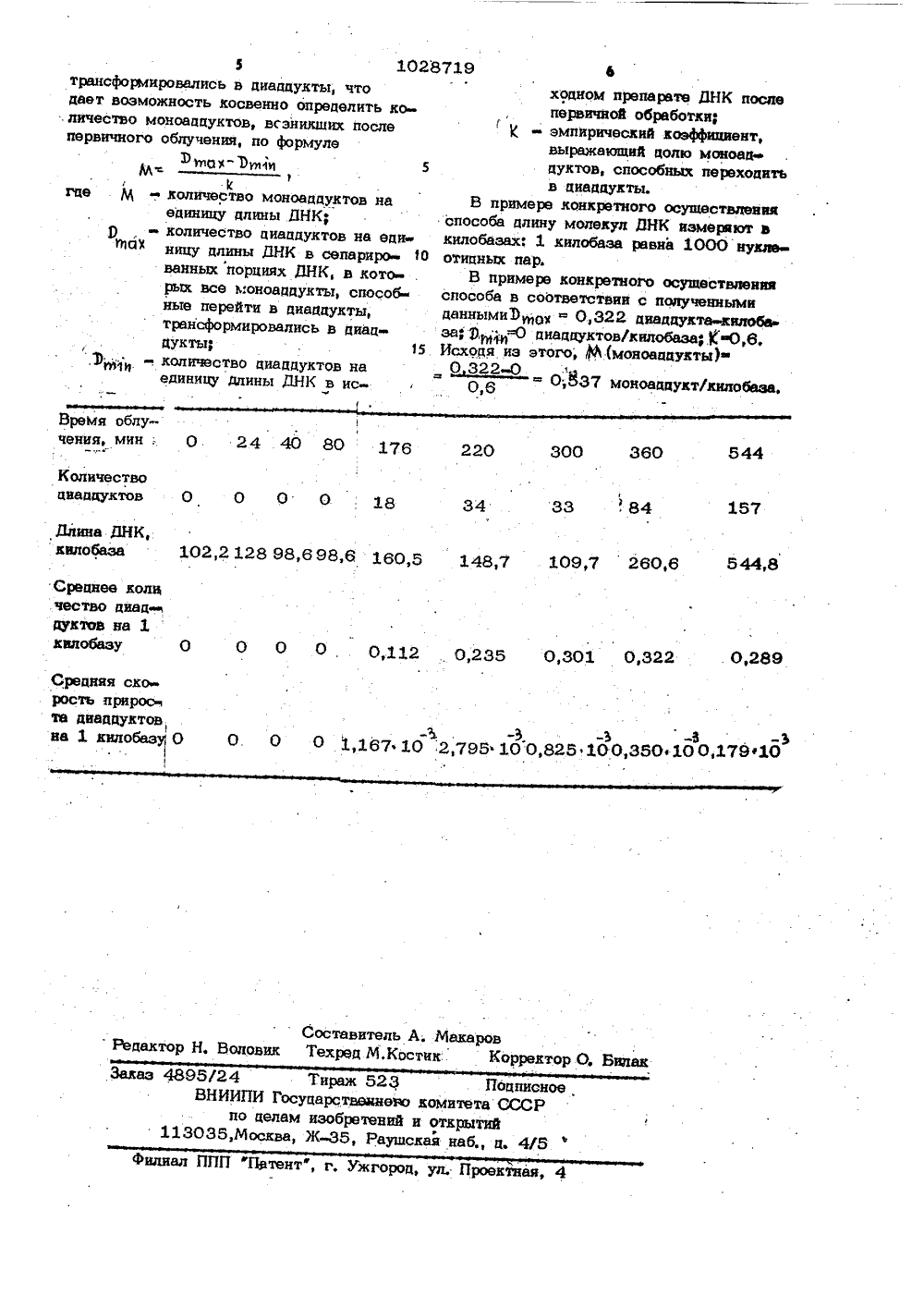
Изобретение относится к способам определения первичных, неферментативных повреждений ДНК, в частности фуро кумариновых моно- и диаддуктов ДНК, . преимущественно 8-метоксипсораленовых, моно- и диаддуктов ДНК.Изобретение может быть использовано в исследованиях по изучению фотохимических процессов связывания молекул ДНК с веществами фурокумари- О нового ряда, для изучения структурно функциональной организации хроматина и хромосом, а также молекулярных механизмов мутагепеза и репарации, Изобретение может быть использовано в прикладных .целях при разработке тест-систем: определение мутагенов окружающей среды, промышленных мутагенов, контроля на мутагенность лекарственных и химичес" ких препаратов. 20Известен способ определения диаддук 3 тов ДНК дпя анализа стурктуры хроматина, согласно которому диаддукты ДНК регистрируют в электронном микроскопе в виде сшивок ДНК Ц . 25Наиболее близким к изобретению является способ определения количества псораленовых моно- и диаддуктов ДНК 23Этот способ основан на выращивании клеток на питательной среде, содержащей 30 р, препаративном выделении ДНК, установлении ее концентрации, инкбации с меченым радиактивным изотопом Ц- триоксаленом, ультразвуковом фрагментировании препарата ДНК, сепарировании массы ДНК на порции, облучении ультрафиолетовыми лучами, разделении хроматографическим способом однонитевых и двунитевых фрагментов ДНК, установлении концентрации ДНК во фракциях и опреде- .4 О пении уровня мечения 11-триоксапеном в тех же фракциях,.Недостатками этого способа являются невозможность получения количественных показателей перехода моноаддуктов в диаддукты ДНК в динамике этого процесса, а также невозможность определения количества моно- и диаддуктов ДНК при разных количественных соотношениях веществ фурокумаринового ряда и молекул ДНК, Кроме того, недостатком этого способа является использование ДНК, предварительно меченой радиоактивным изотопом 2 Р, а также использование меченого М -триоксалена.3Цель изобретения - повышение точности способа путем прямого измерения длины молекул ДНК и количественного подсчета диаддуктов (сшивок) ДНК, а 1028719 2также путем косвенного определенияколичества моноаддуктов,ПНК и скоростиперехода моноаддуктов в диаддукты, упрощение способа путем использования нативной, немеченной радиоактивными изотопами ДНК и немеченного радиоактивными иэотопами вещества фурокумаринового ряда, в частности немеченного 8-метоксипсоралена,Поставленная цель достигается тем,что согласно способу определения моноаддуктов и диаддуктов ДНК, включающему выращивание культуры клеток на питательной среде и препаративное выделение ДНК, установление концентрации ДНК,инкубацию препарата ДНК с веществомфуроКумаринового ряда, сепарацию полученного продукта, облучение его ультра-фиолетовыми лучами, денатурацию егофизическими и химическими средствами,регистрацию диаддукзов, при инкубациипрепарата ДНК с веществом фурокумаринового ряда в качестве препарата ДНКиспользуют нативную ДНК с последующимоблучением ультрафиолетовыми лучами,е/для регистрации моно-диаддуктов испопьэуют электронную микроскопию, а количество моноаддуктов рассчитывают.На чертеже представлен график зависимости количества диаддуктов от времени облучения.П р и м е р . Клетки китайского хомячка выращивают на ййтательной среде по общепринятой методике без внесения в среду веществ, меченных радиоактивными изотопами. Препаративным методом выделяют нативную ДНК и спекгрофотометрически устанавливают ее концентрацию. Всю исходную полученную ДНКинкубируют с веществом фурокумариновогоряда, в частности с 8-метоксипсораленом,не меченным радиоактивными изотопами.Для этого к раствору нативной ДНК вконцентрации 50-280 ь 4 кг/мл добавляютспиртовой раствор 8-метоксипсораленадо конечной концентрации 37 мкг/мл.Инкубацию проводят в 0 Б,0 млбуферного раствора следующего состава: 0,1 М 7 е-НМ, 0,01 И ЭДТА (двунатриевая соль этилдиаминтетрауксуснойкислоты) с рН 8,4 (буферЬ), при 4 фСв течение 18-24 ч.Облучают всю инкубированную с 8 метоксипсораленом ДНК ультрафиолетовъзми лучами с длиной волны 337 нм,с помощью импульсного лазера ЛГИ 21при комнатной температуре. Моуностьдозы составляет 3;9,7 мВт/см, времяоблучения 1 мин.1028 3После облучения ДНК отмываютдважцы путем осаждения ее этиловымспиртом с последующим растворениемв буфере б,Часть исходного обработанного такимобразом препарата ДНК оставляют цлянеобратимой денатурации и электронномикроскопического анализа.Остальную часть препарата ДНК сепарируют на восемь отцельных порций 10обьемом по 100 мкл. Кажцую порцию,ПНКоблучают повторно длинноволновымиультрафиолетовыми лучами с длиной волны 337 нм с помощью импульсного лазера ЛГИнри комнатной температуре 15разное время, в пределах от 20 до 550 мин,Часть исходно обработанной ДНК икаждую порцию повторно-облученной ДНКпо отдельности денатурируют необратимо.для этого ДНК нагревают при 68 С 202 мин в следующем составе; 50 мклраствора ДНК, 10 мкл 30% глиоксаля,100 мкл 99% формамица.После ценатурации готовят известнымспособом препараты для электронной25микроскопии. Электронно-микроскопическим методом измеряют длину молекул ДНК и количество сшивок (циаддуктов) в этих молекулах в каждой порции ДНК, Для этого, при просмотре препаратов в электронном микроскопе фотографируют молекулы ДНК которые содержат оцнонитевые участки в областисшивок однонцтевых молекул ДНК, соответствующие локализации диацдуктов.Измерение количества циаддуктов и35длины молекул ДНК привецены в таблице.Рассчитывают среднее количестводиаддуктов на длину молекул ДНК, выраженную в килобаэах, и среднюю ско 40рость прироста диацдуктов на 1 килоба. зу за период увеличения времени поворотного облучения в двух порциях ДНКи вносят эти данные в.ту же таблицу.Полученные данные длины молекул ДНК 4и количества диаддуктов при разном времени повторного облучения позволяюткосвенно определить среднюю скоростьперехода моноаддуктов в циаццукты эапериод увеличения времени повторногоОблучения в двух порциях ДНК, котораясоответствует тангенсу угла наклонапримой, проходящей через .точки графика,Срецняя скорость перехоца моноадцуктов в диаддукты циап кт/килобазамин. 55, вычисляется по формулеЪ- .У 13,)1 Р с 2 719 4где У, - средняя скорость перехода моноадцуктов в диацдукты;ртангенс. угла наклона прямой;р - количество диаддуктов на единицу цлины молекул ДНК, зарегистрированное в порции ДНКс большим временем повторного облучения;р - количество циаддуктов на еци4ницу цлины молекул ДНК, зарегистрированное в порции ДНКс меньшим временем повторногооблучения;- время повторного облученияЯпорции ДНК с более длительнымоблучением;ь- время повторного облученияпорции ДНК с менее цлительным облучением.В примере конкретного осуществленияспособа средняя скорость прироста диацдуктов на ециницу длины ДНК в цвух порциях повторно обработанной ДНК, вычисленная по указанной выше формуле; прец"ставлена следующими цифровыми данными.Срецняя скорость прироста диацдуктовна 1 килобазу (на 1000 .нуклеотидныхпослецовательностей) за время облучения0 - 24; 24 - 40; 40 - 80 мин равна 0;за время облучения, мин равна соответственно 80 - 176, - 1,167 10176 - 220 - 2,795 10 ; 220 - 3000,825" 10 , 300 - 360 - 0,35010- 360 - 544 - 0,179:10" ,Вычисляют количество моноацдуктов,возникавших при первичном облучении исхоцного продукта ДНК косвенным методом.В трм случае, когда после облучения цлинноволновыми ультрафиолетовыми лу чами вешество фурокумаринового ряда удаляют из среды, содержащей ДНК, диац цукты при повторном облучении длинно- волновыми ультрафиолетовыми лучами возникают только из первично индуцированных моноацдуктов ДНК. В порциях повторно обработанной ДНК при увеличении времени облучения длинноволновыми ультрафиолетовыми лучами количество первоначально возникших моноаццуктов уменьшается за счет образования циаццуктов, При этом возникает такая ситуация, при которой количество диацдуктов в некоторых пор циях ДНК в расчете на единицу цлины молекул ДНК не возрастает, несмотря на увеличение времени облучения цлинноволновыми ультрафиолетовыми лучами. Это свицетельствует о том, -что все моноаддукты, способные образовать циаацукты,1028719 Время облучения, мин:. О 24 40 80 176 844 300 360 220 Количествоциацауктов О О 0 0 18 33 84 34 Длина ЛНК,кило база 102,2 128 98,6 986 160,8 148,7 1097 6 Срецнее количестно диацдуктое на 1килобазу 0 О О 0 0112 О 238 0301 О 322 0,289 Средняя скорость прироста диаддуктовна 1 килоО-Ъ . -3. -Ь -3 . -Ъ1167 10 2,798 1 ОО 828 10038010017910 Составитель А. МакаровРедактор Н. Воловик Техред М,Костик: Корректор О, Билак Заказ 4898/24 Тираж 82 Подписное,ВНИИПИ Госуцарстищнвэо комитета СССР по целам изобретений и открытий 113038,Москва, Ж, Раушская наб., д. 4/8Филиал ППП "Патент", г, Ужгород, ул. Проектная, 4 5трансформировались в диаддукты, что дает воэможность косвенно определить ко. личество моноадцуктов, всэникших после первичного облучения, по формулеион- ЭеЙ 5Мгце М . количество моноаддуктов наединицу длины ДНК;р - количество циацдуктов на еди"ИО" ницу длины ДНК в сепариро 30ванных порциях ДНК, в которых все моноаддукты способные перейти в диадцукты,трансформировались в диацдукты 15,Э ,; -. количество диаддуктов наединицу дпнны ДНК в иск 6хоаном препарате ЛНК послепервичной обработки- эмпирический коэффициент,выражающий целю моноаддуктов, способных перехоцитьв циаааукты.В примере конкретного осуществления способа длину молекул ДНК измеряют в кнлобазах: 1 килобаза равна 1000 нуклвотицных пар.В примере конкретного осуществления способа в соответствии с полученными даннымиЭп,о = 0,322 диацдукта-.иилоба зади"0 цнацауктов/килобаза,К 0,6, Исходя из этого, М(моноаццукты) О 322-0- - О;53 Т моиаук/нпобаа.
СмотретьЗаявка
3262236, 29.12.1980
ИНСТИТУТ ОБЩЕЙ ГЕНЕТИКИ АН СССР
ЕВГРАФОВ ОЛЕГ ВАДИМОВИЧ, МАКЕДОНОВ ГЕННАДИЙ ПЕТРОВИЧ, ФЕДОТОВ АЛЕКСАНДР РАДИЕВИЧ, ТИНЯКОВ ЮРИЙ ГЕОРГИЕВИЧ
МПК / Метки
МПК: C12N 15/00
Метки: диаддуктов, днк, моноаддуктов
Опубликовано: 15.07.1983
Код ссылки
<a href="https://patents.su/4-1028719-sposob-opredeleniya-monoadduktov-i-diadduktov-dnk.html" target="_blank" rel="follow" title="База патентов СССР">Способ определения моноаддуктов и диаддуктов днк</a>
Установка для облучения семян гамма-лучами

Номер патента: 130261
Опубликовано: 01.01.1960
Авторы: Аракелов, Бибергаль, Дещинский, Коротков
Метки: гамма-лучами, облучения, семян
...цилиндрическая полость облучения 2; камеры 3 для облучаемого материала (семян) желобчатой формы; цилиндрический шток 4, объединяющий камеры в общий стержень при помощи защитных сэанцовых отрезков 5 и передвигаемой в горизонтальной цилиндрической выемке защитного корпусаустановки б.Подлежащие обработке гамма-лучами порции семян, помещаемые в камеры 3 поочередно вводятся в полость облучения 2 и подвергаются там воздействию гамма-лучей, поступающих из облучателей 1. По скончании облучения соответствующая семенная камера выдвигается из полости облучения 2, в то время как другая камера, загруженная зерном, вэодится в полость облучения. В момент, когда одна камера разгружаетсяпроизводиться загрузка другой камеры.У .ЗО 26 Проникновение...
Способ купирования алкогольного абстинентного синдрома

Номер патента: 1135476
Опубликовано: 23.01.1985
Авторы: Береснев, Беспалов, Лях, Слабунов, Сосин, Худенский
МПК: A61K 35/14
Метки: абстинентного, алкогольного, купирования, синдрома
...резкий запах перегара. Язык обложен грязно-серым налетом. Кожныепокровы влажные. Тахикардия - 110 ударов вмин. АД - 150/90 мм рт. ст,Тремор всего тела, неустойчивостьв позе Ромберга. Сухожильные рефлексы с рук и ног с расширенными зонамивызывания, симметричные Нистагмоидные подергивания глазных яблок привзгляде в стороны. Речевому контактудоступен, ориентирован в месте, времени и собственной личности. Напряжен,тревожен, настроение снижено, выражение лица страдальческое. В беседе быстро истощается, периодически на глазах слезы, просит о помощи,Для полной нормализации состояния после запоя, как явствует иэ меддокументации, объективных сведений родственников и сообщения больного, ему в среднем требуется около двух недель. Сразу после...
Строительный гелиокомплекс

Номер патента: 912862
Опубликовано: 15.03.1982
Авторы: Баланюк, Мелуа, Селиванов
МПК: E04B 1/34
Метки: гелиокомплекс, строительный
...силам от, враще.пия экрана 3 вокруг вертикальной оси 8 атакже для улучшенного восприятия несиммет.ричных динамических ветровых нагрузок. Поэтому виток спирали 14 представляет собоюувеличенную в диаметре проекцию на цилиндрическую поверхность образующей спиралиопорного стакана 10. Ближайшие друг к другуконцы звеньев 9 соединены стержнем 16, на 50котором между упорами 17 шарнирно насажен. стакан 18 консольной опоры 19 соединенньйсиловым, например, вертикальным бесконечнымтросом 20, пропушенным через привод известного типа (не показан). 55Для вращения вокруг горизонтальной оси 6,совпадающей с осью стержня 6 экран 3 оборудован опорами 21, к которым подведен трос 422, переброшенный затем через ролик 23, оськоторого жестко соединена с...
Синхронизатор для растрового дисплея

Номер патента: 1606993
Опубликовано: 15.11.1990
МПК: G09G 1/16
Метки: дисплея, растрового, синхронизатор
...счетчики 1 и 5 соответственно первого 4 и второго 8 блоков пересчитывают входные импульсы до периода соответственно строчной и кадровой развертокНа выходе счетчиков 1 и 5 Формируются импульсы с периодом соответствующей развертки и длительностью, равной периоду входных импульсов. Эти импульсы являются запускающими сигналами для вторых и третьих счетчиков соответствующих блоков.Коэффициенты пересчета вторых счетчиков 2 и 6 соответственно первого 4 и второго 8 блоков определяют момент начала соответственно строчного и кадрового синхроимпульсов. Изменяя эти коэффициенты, можно изменять положение изображения .на экране дисплея в горизонтальном и вертикальном направлениях.Коэффициенты пересчета третьих счетчиков 3 и 7 первого 4...
Способ определения свободного диоксида кремния в угольной и породной пыли

Номер патента: 1357843
Опубликовано: 07.12.1987
Автор: Федорова
МПК: G01N 31/22
Метки: диоксида, кремния, породной, пыли, свободного, угольной
...С для испарения жидкости, после чего помещают на 7 мин в муфельную пеныонагретую до 850 С. После охлаждения в тигель вносят 10 мл 5%-ного раствора углекислого натрия и нагревают до полного растворения плава. Фильтруют через зольный фильтр в мерную колбу емкостью 50 мл. Тигель промывают 3 раза водой при нагревании и промывшие воды сливают через тот же фильтр. Раствор нейтрализуют 7 мл 1 н. Серной кислоты и по охлаждении доводят объем до 50 мл.Для анализа берут 2 мл этого раствора, разбавляют до 20 мл, приливают 1 мл 7,5%-ного чистого серно-"кислого раствора молибдата аммония и через 5 мин 5 мл 5%-ного раствора лимонной кислоты и 5 мл. 10 н. серной кислоты. Облучение раствора проводят в течение 5 мин- ультрафиолетовым Осветителем ВИОв...
Предыдущий патент: Штамм 97 фага наг-вибрионов 0-41 серовара
Следующий патент: Штамм инми-кз-2, используемый для очистки сточных вод от 2 хлорбензойной кислоты
Случайный патент: Бесконтактная электрическая машина