Способ получения аналогов тетраплоидной ржи
Похожие патенты | МПК / Метки | Текст | Заявка | Код ссылки
Номер патента: 1807842
Автор: Титаренко





Текст
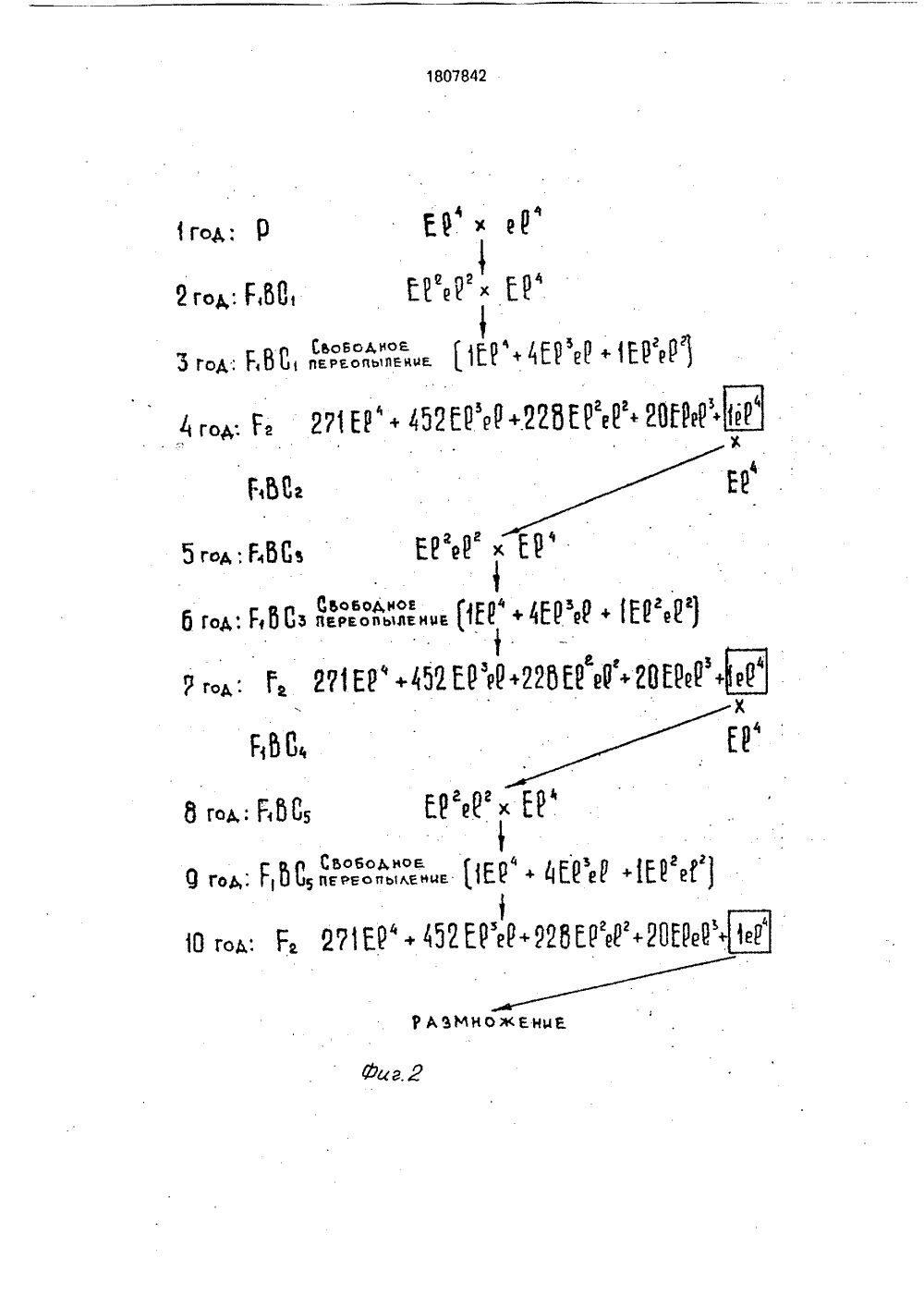
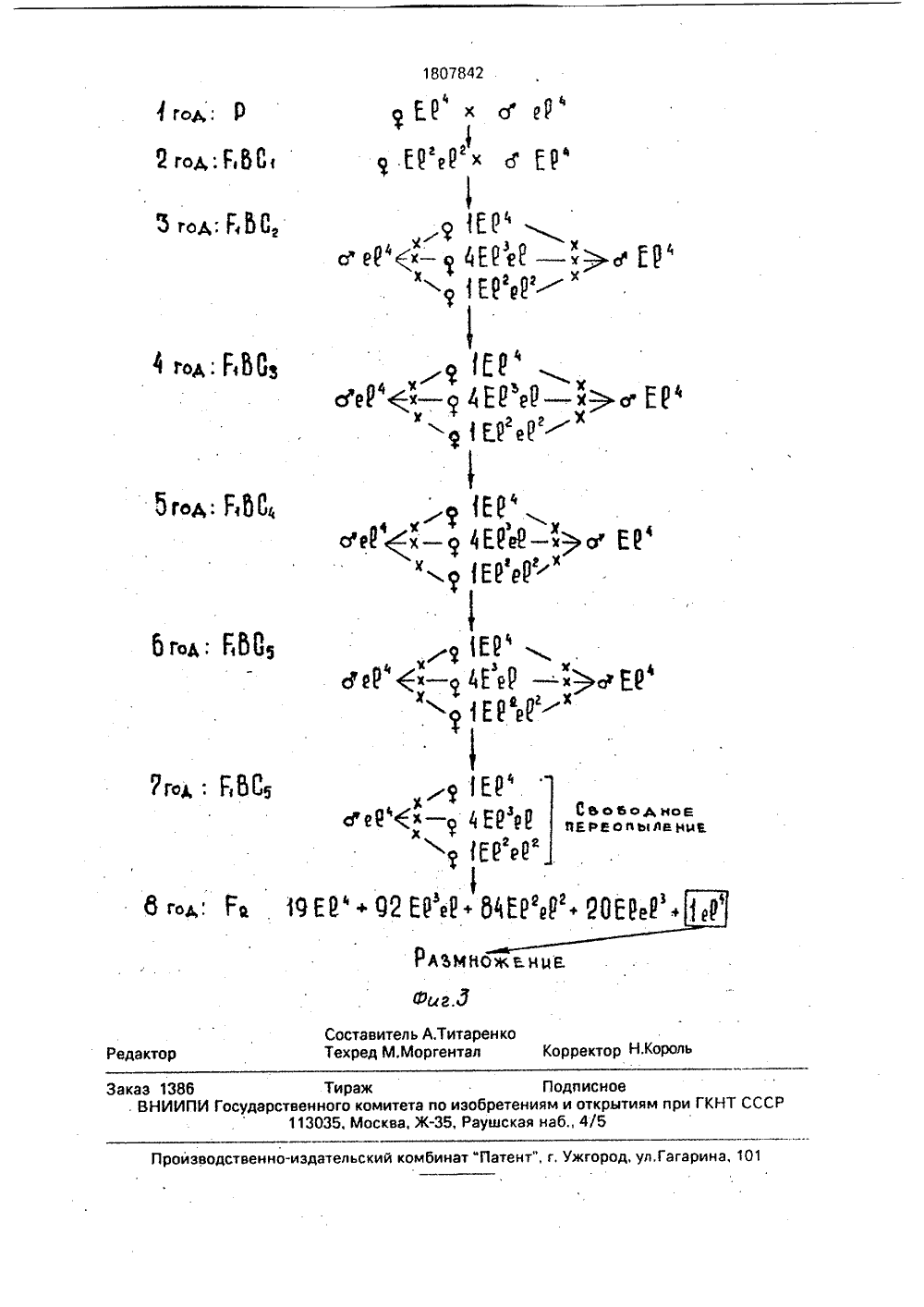
СОЮЗ СОВЕТСКИСОЦИАЛИСТИЧЕСРЕСПУБЛИК 07842 А 9) и 1) Н 1 04 51) ГОСУДАРСТВЕННОЕ ПАТЕНТНОВЕДОМСТВО СССР(ГОСПАТЕНТ СССР) ИЕ ИЗОБРЕТЕНИ О ПАТЕ Н сновы од ред,ОО Изобретение относится к сельскому хо- и может быть использовано в селекции этйству, в частности к селекции озимой ржи, и других растений.. Краснюк А.А. Некоторые данные по генетике ржи, - Селекция и семеноводство, 1936, М9, с. 50-53.Федоров В.С, идр, Генетика ржи, - Генетика; 1970, т,6, М 5, с 5-14,Гуляев Г,В, и др., Использование ограниченного размера популяции в селекции перекрестноопыляющихся культур, ИзвестияТСХА, 1973, выг), 4 с. 74-79,(54) СПОСОБ ПОЛУЧЕНИЯ АНАЛОГОВ ТЕТРАПЛОИДНОЙ РЖИ(57) Использование; сельское хозяйство, способы генетического отбора и получения аналогов тетраплоидной ржи. Сущность изобретения: способ включает скрещивание материнской формы с отцовской формой-носителем необходимого признака, получение гибридов Р 1, возвратные скрещивания и интенсификацию необходимых генотипов. Используют отцовскую форму, обладающую рецессивным геном безлигульности и параллельно с возвратными проводят анализирующие скрещивания на всех генотипах Р 1 ВС 1-и. По результатам анализа потомств анализирующих скрещиваний выделяют гетерозиготные генотипы, по которым отбирают гибриды, полученные от возвратных скрещиваний для последующего цикла возвратных и анализирующих скрещиваний, В качестве аналога идентифицируют безлигульные генотипы Г 2 по- ф следнего возвратного скрещивания.Выделение гетерозиготных генотипов проводят в фазе третьего листа - колошение, 1 ь з,п. ф-лы, 3 ил., 1 табл, 00Целью изобретения является ускорение процесса получения безлигульного аналога.Поставленная цель достигается тем, что в качестве отцовской используют форму, обладающую рецессивным геном безлигульности, параллельно с возвратными проводят анализирующие скрещивания на всех генотипах Г ВС-, по результатам анализа потомств анализирующих скрещиваний выделяют гетерозиготные генотипы, по которым отбирают гибриды, полученные от возвратных скрещиваний для последующего цикла возвратных и анализирующих скрещиваний, а в качестве аналога идентифицируют безлигульные генотипы в Р 2 последнего возвратного скрещивания. Выделение гетерозиготных генотипов проводят в фазе 3 листа - колошение.Сопоставительный анализ заявляемого технического решения с прототипом показывает, что заявляемый способ отличается от известного тем, что получение аналога ведется по рецессивному гену, фенотипически не проявляющегося в Г 1 до цветения, поэтому параллельно с возвратными проводят анализирующие скрещивания для выделения в фазе 3 листа - колошение гетерозиготных генотипов и полученных с ними гибридов для последующего возвратного скрещивания, в качестве аналога идентифицируют безлигульные генотипы в Р 2 последнего возвратного скрещивания.Изучение патентной и научно-техниче, ской литературы свидетельствует о том, что сочетание возвратных и анализирующих скрещиваний, способствующих проведению непрерывного скрещивания при ускоренномполучении аналогов, различающихся по рецессивному гену, является новым и в селекционной работе с полиплоидами не применялось.Предлагаемый способ предусматривает использование известных данных, Известно, что в случаях, когда передаваемый признак обусловлен рецессивным геном, т.е, фенотипически не проявляется в Г можно проводить 2-3 возвратных скрещивания подряд, затем выращивать потомство от второго или третьего беккроссов и в нем отбирать гомозиготные по рецессивному гену растения.Такой путь получения аналогов сокращает период их получения до 9-10 лет, при большем объеме гибридизации. Однако, если для диплоидной ржи можно проводить три скрещивания подряд, то для аутотетраплоидной, в связи с более сложным характером расщепления, только два, причем концентрация необходимых генотипов будет невысокая, Аналог может быть полученчерез 10 лет,Известно также, что в случаях, когда передаваемый признак фенотипически не5 проявляется в Р 1 до цветения, используютсяупреждающие (возвратные) скрещивания спараллельным самоопылением части генотипов, включенных в гибридизацию, для ихиндентификации в потомстве, Это позволя"0 ет получить аналог через 8 лет, т,е, в такиеже сроки, как и при непрерывном беккроссес источником доминантного признака.Данный способ разработан и применимдля самоопылителей, у которых при само 155 опылении завязывается достаточное количество семян для идентификации генотиповв потомстве. У перекрестников же завязываемость зерен при самоопылении низкая, зависит от многих факторов и составляет дляржи 0,02-3,0 , в среднем по данным многихавторов менее 5 , Поэтому использованиесамоопыления для идентификации генотипов у ржи, тем более у аутотетраплоидной,неприемлемо по той причине, что большая25 часть генотипов не будет идентифицирована.Известен способ получения изогенныхлиний по рецессивному признаку семени уподсолнечника, предусматривающий скре 30 щивание рецидиента с донором рецессивного признака, полученный гибрид накаждом этапе беккросса используется в качестве отцовского компонента, Для дальнейшего насыщения отбираются гибридные35 семена от парных скрещиваний, в которыхотцовской формой были гетерозиготныерастения, индентифицированные по расщеплению в потомстве от самоопыления.Недостаток данного способа в том, что40 наряду с самоопылением, контроль гетерозиготных генотипов осуществляется по отцовской линии. Если это применимо дляподсолнечника при отсутствии кущения, тодля ржи данный способ контроля гетерози 45 готных генотипов усложняет процесс, получения аналогов,Идентификация генотипов проводитсяобычно путем анализа расщепления потомств от самоопыления или анализирую 50 щих скрещиваний, т.е. скрещивания Е 1 сгомозиготной по рецессивному гену формой. У ржи анализирующие скрещиванияприменяются для выделения гомозиготныхпо доминантному гену короткостебельно 55 сти генотипов как на диплоидном, так и на тетраплоидном уровне,Известно, что наследование признакову аутотетраплоидной ржи более сложное, чем у диплоидной. Обусловлен тем, что удиплоидов при двух аллелях образуется триразличных генотипа, тогда как у аутотетраплоидов пять, в связи с чем и более сложныйпроцесс их идентификация, В зависимостиот типа расщепления и способа идентификации, частота фенотипов в потомстве может быть различна (смтаблицу). Дляидентификации одного и того же генотипа,в большинстве случаев дуплекса, при анализирующем скрещивании потребуется значительно меньший объем выборки, чем при 10самоопылении,Хромосомный или хроматидный типырасщепления крайние варианты расщепления у аутотетраплоидов и в чистом виде,встречаются редко . Нами для расчета обьема и 15схемы работы взят хромосомный тип расщепления, так как суть при этом не меняется, а достоверность выделениягетерозиготы будет выше, потому что длядоказательства этого типа расщепления 20объем выборки должен быть больше, чемдля хроматидного типа расщепления.Безлигульность у ржи рецессивныйпризнак, расщепление в Г 2 идет по,простоймоногибридной схеме 3:1. Полученные авторами результаты показывают, что лигульные и безлигульные растения можноразличать уже в фазе трех листьев,Таким образом, авторами обнаруженоновое свойство уже известных признаков, а 30именно: непрерывные возвратные (упреждающие) скрещивания позволяют быстрополучить безлигульные аналоги, различающиеся по рецессивному гену, за счет постоянного использования в гибридизации 35гетерозиготных по гену е генотипов, идентифицированных анализирующими скрещиваниями в фазе 3 листьев.У гибридов аутотетраплоидной ржи гетерозиготы представлены триплексами и 40дуплексами (фиг. 3). Использование самоопыления для идентификации этих генотипов неэффективно, поскольку триплекс прихромосомном расщеплении не дает расщепления, а при хроматидном - необходима выборка, которую не получить отсамоопыления отдельных генотипов. Дляидентификации дуплексов потребуется неменее 80-140 зерен, т,е. исходный генотипдолжен иметь, при завязываемости зерен от 50самоопыления, показанной выше, с учетомполевой всхожести, 40-50 продуктивных побегов, что для тетраплоидов, отличающихсяпониженной кустистостью, нереально,Достоверная идентификация генотипов 55у тетраплоидов может быть достигнута использованием анализирующих скрещиваний. Так, для идентификации того жедуплекса потребуется всего 20-25 зерен, которые легко получить от опыления двух кастрированных колосьев (завязываемость зерна при гибридизации в среднем составляет 40 ), К тому же в случае хроматидного расщепления при анализирующем скрещивании могут быть идентифицированы даже триплексы.Безлигульный аналог тетраплоидной ржи может быть получен обычным путем (фиг. 1) через 13 лет, при этом обьем гибридизации в годы скрещивания составляет 25- 30 колосьев (из расчета 1 колос с растения, учитывая то, что минимальное число растений, отражающих популяцию у ржи составляет 25), На изоляторах проводится размножение Р и выделение в Р 2 безлигульных растений, концентрация которых составляет при хромосомном типе расщепления 2,8 , при хроматидном - 4,6/.Использование второго подряд скрещивания, т.е. получение ВС 1, позволяет сократить срок получения аналога до 10 лет (фиг. 2). Объем гибридизации и потребность в изоляторах остается на уровне обычной схемы, но концентрация безлигульных растений в Г 2 составляет 0,1 при хромосомном и 0,3 при хроматидном типах расщепления.Предлагаемый способ позволяет получить безлигульный аналог через 8 лет, аналогично схеме беккросса с источником доминантного признака, при этом первые два года объем гибридизации остается такое же, как и в предыдущих схемах (фиг, 3), На 3-6 годы увеличивается число генотипов, используемых за материнскую форму до 6 и параллельно с возвратными проводятся анализирующие скрещивания, поэтому ежегодный объем гибридизации увеличивается до 450 колосьев из расчета 2 колоса с растения для анализирующего и 1 колос для возвратного скрещивания.На 7 год проводятся только анализирующие скрещивания с целью выделения гетерозиготных генотипов, в частности дуплексов, для последующего перепыления их в Г 2. Число генотипов Р 2 рассчитано с учетом перепыления дуплекса с другими генотипами в Г 1, Концентрация безлигульных растений в Г 2 при хромосомном типе расщепления составляет 0,5 , при хроматидном - 1,3 .Таким образом, на основании анализа известных решений предлагаемое техническое решение обладает существенными отличиями.При этом получен положительный эфР фект - предложенный способ позволяет получить безлегульный аналог тетраплоидной ржи за более короткий срок (фиг, 3).1807842троль за генотипами, По результатам анализа расщепления потомств анализирующих скрещиваний проводят выделение гетероэиготных генотипов и по ним отбирают гибриды, полученные от скрещивания гетерозиготных генотипов с рекуррентом для последующего цикла возвратных и анализирующих скрещиваний, Остальные потомства бракуются.После получения аналога ВСв отбираютгибрит от скрещивания гетероэигот с рекуррентом для переопыления, дающий расщепление 1:4;1. Проводятся анализирующие скрещивания для выделения дуплексов, Все генотипы Е 1 ВСв свободно переопыляются между собойВысевают Ег переопыленных дуплексови выделяют в фазе 3 листьев безлигульные тетраплоидные растения.Формула изобретения1. Способ получения аналогов тетраплоидной ржи, включающий скрещивание материнской формы с отцовской формой - носителем необходимого признака, получение гибридов Е 1, возвратные скрещивания и идентификацию необходимых генотипов, о т л и ч а ю щ и й с я тем, что, в качестве отцовской используют форму, обладающую рецессивным геном безлигульности, параллельно с возвратными проводят анализирующие скрещивания на всех генотипах Е ВС 1-, по результатам анализа потомств анализирующих скрещиваний выделяют гетерозиготные генотипы, по которым отбирают гибриды, полученные от возвратных скрещиваний для последующего цикла возвратных и анализирующих скрещиваний, а в качестве аналога идентифицируют безлигульные генотипы в Ег последнего возвратного скрещивания.2. Способ по и. 1, о т л и ч а ю щ и й с ятем, что выделение гетерозиготных генотипов проводят в фазе третьего листа - колошение,Отношения расщепления по одному гену (А: а ) на аутотетраплоидном уровне1 ьоьодноаРЕООьАВ иР Р + б 151ЬОй ОЕЕ ойъАЕиц еР еР+ 1 б ЕЕ Сьаьодйов ЕРЕ ОПЫЛЕНИИ Е) е 8 е 0 ВИе 1( ег+ввиЦ . Р Го бг д:ГЬС ЕРе(х Е(ц Е(цЕ(е( Р 0 ЬО 60 ДнОегьд. ьОВ пгьеьльлен еР О, Р, КЕР.452 ЕРъР 22 ВЕРе( 20 ЭМНОЖЕН 44 В:г, ег 1 н ахи еевгвг.еаеЯХЬМе Н,уеР" ,- - 4(еРбЕР1 йГ 04 Б 815 ,. 1 И"с Р (1 Г 1 11 пеРеользлени%.Юе 16 год. Га Й Е" + 02 НЙ+ 61 НИ+ 205 Мф Д 2 ГО:Г И 1 бгод: ЕЬО 1 АЪМНОжа них Фиг,5 Составитель А.ТитаренкоТехред М.Моргентал Корректор Н,Король Редактор Производственно-издательский комбинат "Патент", г. Ужгород, ул,Гагарина, 101 Заказ 1386 Тираж Подписное . ВНИИПИ Государственного комитета по изобретениям и открытиям при ГКНТ СССР 113035, Москва, Ж, Раущская наб., 4/5
СмотретьЗаявка
4935019, 06.05.1991
НАУЧНО-ИССЛЕДОВАТЕЛЬСКИЙ ИНСТИТУТ СЕЛЬСКОГО ХОЗЯЙСТВА ЦЕНТРАЛЬНО-ЧЕРНОЗЕМНОЙ ПОЛОСЫ ИМ. В. В. ДОКУЧАЕВА
ТИТАРЕНКО АЛЕКСЕЙ ВАСИЛЬЕВИЧ, ТИТАРЕНКО ЛИДИЯ ПЕТРОВНА
МПК / Метки
МПК: A01H 1/04
Метки: аналогов, ржи, тетраплоидной
Опубликовано: 07.04.1993
Код ссылки
<a href="https://patents.su/7-1807842-sposob-polucheniya-analogov-tetraploidnojj-rzhi.html" target="_blank" rel="follow" title="База патентов СССР">Способ получения аналогов тетраплоидной ржи</a>
Способ отбора генотипов зерновых культур, устойчивых к насекомым lема меlаnорus l

Номер патента: 1449063
Опубликовано: 07.01.1989
Автор: Жогин
МПК: A01H 1/04
Метки: lема, генотипов, зерновых, культур, меlаnорus, насекомым, отбора, устойчивых
...популяции используют растения с признаком свернутый лист (СЛ), контролируемым рецессивным геном.Рацессивный генетический маркер свернутый лист индуцируется у тритикале с помощью химических супермутагенов. У других культур (пшеницы, ржи, ячменя, овса) также получают подобные мутации согласно закону гомологических рядов Н. И. Вавилова или же передают путем скрещивания СЛ-тритикале с пшеницей, рожью и ячменем.Пример. Проводят обработку семян озимого тритикале КМ 4349 раствором химического супермутагена, затем их высушивают в течение 32 ч. Обработанные таким образом семена высевают в поле и выращивают растения М. Убирают комбайном.Осенью сеют полученные семена (с нормой 4 млн. всхожих зерен на 1 га), выращивают растения Мг. Весной производят...
Способ селекции аутотетраплоидных популяций ржи

Номер патента: 1777723
Опубликовано: 30.11.1992
Автор: Титаренко
МПК: A01H 1/04
Метки: аутотетраплоидных, популяций, ржи, селекции
...поколения второго анализирующего скрещивания в одноименных по изучаемому генотипу группах. Сохранение генотипов проводят путем их инцухтирования,Способ осущезом.Высевают семьи аутотетраплоидной популяции в массиве опылителя - источника рецессивной гомозиготы, совпадающего по срокам цветения, Расщепляющиеся семьи весной бракуют, а на растениях нерасщепляющихся семей одновременно проводят инцухтирование 5 - 6 колосьев для сохранения генотипов и первые анализирующие скрещивания.Высевают первое поколение анализирующего скрещивания в массиве опылителя ствляют следующим обра,Стенин аэ 4142 ВНИИП Тираж Подписноедарственного комитета по изобретениям и открытиям при ГКНТ С113035, Москва, Ж, Раушская наб., 4/5 изводственно-издательский комбинат...
Способ селекции аналогов-восстановителей фертильности линий кукурузы

Номер патента: 1092771
Опубликовано: 23.04.1985
Автор: Гонтаровский
МПК: A01H 1/04
Метки: аналогов-восстановителей, кукурузы, линий, селекции, фертильности
...восстановительная способность этих аналогов в отдельных гибридных комбинациях, Ген КГ не у всех генотипов обеспечивает необходимую для нормального опыления фертнльность, что является причиной череззерницы початков и снижения урожая зерна кукурузы.Цель изобретения " создание надежных аналогов-восстановителей фер 55 тильности с высокой Фертильностью и восстановительной способностью.Поставленная цель достигается тем, что в процессе насыщающих скрещиваний проводятдополнительно скрещивание,линии с линией - источником генаКГ , а выделение генотипов провочагдят по наличию в них обоих геновчагКГ и КГ в гомозиготном состоячагнии. Ген КГ является геном-восста.новителем, обладающим более слабымдействием, чем ген КГ . Оба гена,находясь в.одном...
Способ создания доноров раздельноцветковости у сахарной свеклы

Номер патента: 1575330
Опубликовано: 30.10.1993
Авторы: Вепрев, Малецкий, Шавруков
МПК: A01H 1/04
Метки: доноров, раздельноцветковости, сахарной, свеклы, создания
...Е 1 будет сростноцветковым. Такие линии являются специфическими РЦ аллелям.. Выделяют линии, которые при скрещивании с другими линиями будут иметь все потомство Р 1 раздельноцветковым, Такие линии являются универсальными РУ анализаторами.4Скрещивают любое СЦ растение с универсальным РЦ анализатором, т.е. с линией гомозиготной по аллели т 5 щб. Это позволяет выделить любые РЦ аллели, В Р 1 выделяют РЦ растения, которые и являются донорами раздельноцветковости.Для точной идентификации РЦ аллелей у любого РЦ растения выделенные доноры дополнительно последовательно скрещивают с линиями специфическими анализаторами гомозиготными по аллелям пиа 2, ПЗ 1 ПЗт ГП 4 П 14,РЦ аллель любого СЦ растения будет идентичен РЦ аллелю этого специфического...
Способ отбора продуктивных генотипов злаковых растений
Номер патента: 1262763
Опубликовано: 27.03.2000
Автор: Малюженец
МПК: A01H 1/04
Метки: генотипов, злаковых, отбора, продуктивных, растений
Способ отбора продуктивных генотипов злаковых растений путем поярусного измерения ассимиляционной поверхности и выделения генотипов с максимальным значением данного показателя, отличающийся тем, что, с целью повышения точности отбора и общей продуктивности отобранных генотипов, для возделывания отбираемых генотипов в условиях богарного земледелия выделяют генотипы с максимальной ассимиляционной поверхностью в средней части растения, а при возделывании в условиях орошаемого земледелия - с максимальной ассимиляционной поверхностью в нижней части растения.
Предыдущий патент: Способ отбора элитных растений льна-долгунца
Следующий патент: Способ микроразмножения шафрана посевного (crocus sатivus l. )
Случайный патент: Приспособление для поворачивания лодки при постройке